科学
无标题无名氏No.58087530
2023-06-14(三)22:12:20
ID: plwvFwS 回应
四维空间中,小学二年级数学王子秦老师正在准备明早的四维胞几何课。
王老师已经从数学组借来了四次方胞教具和球胞教具。
四次方胞有八个体,每个体都是同样大小的立方体,这立方体学生们已经在平体几何学过了:答上来立方体有六个面,每个面都是同样大小的平方形奖励一块玉米糖吧。哈哈,要是有五维空间的外星人,那这道题还能再嵌套一层,五次方超胞有十个胞,每个胞都是同样大小的四次方胞……王老师心想。
球胞只有一个体……不对,这话不能讲,会混淆孩子们对于表体的定义。小学生只用明白平体就好,曲体上大学再研究。球胞是到球心距离相等的点的集合,这个距离叫半径。体积二分之π方r四次方,这个公式初中还是高中才讲吧。
球柱和圆柱柱的教具数学组没有,不借了,看课本图片应该都懂。分不清这俩东西就背口诀呗,什么“三圆一方”,“两圆两方”。
双圆柱是这节课的难点,考纲不考,但很常用,得讲。高中老师讲起来倒方便,列方程就行了:x²+y²≤1, z²+w²≤1. 小学要怎么讲呢?王老师决定自己做一个教具。他打开4D打印机,w=-1层是一个高度为0,半径为1的圆柱;w=-0.8层还是圆柱,半径不变,高度为0.6... 一直到w=1层,打印完毕。
第二天数学课上,小明,小红,小刚围坐在球桌边,观察着双圆柱。请问,关于这个几何胞,他们会说些什么?
回应有 4 篇被省略。要阅读所有回应请按下回应链接。
无标题无名氏No.58389049
2023-07-01(六)14:12:36
ID: yyCsokr
不对说错了
无标题无名氏No.58389222
2023-07-01(六)14:27:05
ID: yyCsokr
应该是类似一个坡型
无标题无名氏No.58400699
2023-07-02(日)09:49:22
ID: B3EjIit
(>д<)老师这是什么
无标题无名氏No.58586719
2023-07-13(四)12:56:41
ID: dqzcIpu
好!再来一个(`ヮ´ )
无标题无名氏No.58587248
2023-07-13(四)13:27:32
ID: 3XzWfhs
秦老师(ᯣ ̶̵̵̵̶̶̶̶̵̫̋̋̅̅̅ᯣ )
无标题无名氏No.58333398
2023-06-27(二)16:55:15
ID: 4dXfwrk 回应
子宫切除这样的手术会对阴道和腹腔做什么处理?虽然反复看到了很多讨论(和争吵)但是阴道会留着吗,( ゚∀。)7会加入人造支撑模拟原有空间布局吗,其他器官切除会吗(比如肾)
声名:提问源于好奇,灵感来自外国去性别化手术(会在腿之间只留两个洞的那种平坦形式的),如果可行性大就是一个如何在土里种树的问题,如果可行性小就是一个如何在太空种树的问题,不可行就是如何在异次元种树的问题。
无标题无名氏No.58333458
2023-06-27(二)16:59:53
ID: UadYykj
上一次见到子宫切除手术还是在医学案例分析,说到有时候生孩子子宫出血量太大,没办法止住,最后一招就只能是子宫切除,来勉强把命保住
固定子宫的话有子宫圆韧带等结构,固定阴道的话有盆底的一些结构,总之固定二者的东西不太一样。
我是一个刚考完试脑子不太清醒的医学肥(;´Д`)
无标题无名氏No.58587106
2023-07-13(四)13:19:25
ID: 1gA5Yrd
其他不清楚,阴道肯定会留着的呀……只是切除子宫,有的病切除子宫 宫颈也会问你保留还是切除的。(并且子宫实际上也没有很大)…而且阴道只是一个通道…怎么切除…就算切除了一层还是有洞啊…没必要…而且听着都疼|д` )也不影响生活
无标题无名氏No.57823526
2023-06-02(五)12:17:59
ID: plwvFwS 回应
脑洞一个外星生物。如果傅里叶变换对这种生物来说是一种类似于加减乘除的基础运算,那么它们的数学和科学体系会有什么变化?
回应有 5 篇被省略。要阅读所有回应请按下回应链接。
无标题无名氏No.57827316
2023-06-02(五)15:58:51
ID: ks4hhRF
那他们就很难算我们的加减法( ゚∀。)
无标题无名氏No.57828464
2023-06-02(五)16:57:04
ID: LnKu7RG
>>No.57823783
人算加减乘除本质上是查表,先记住十以内加减法,再记住进位退位规则,再记住乘法表和除法规则,然后加减乘除就都可以算了。剩下乘方开方对数指数三角函数积分微分还是靠记住规则和几个常用值,然后化简成简单运算来算。
从傅立叶变换开始感觉也就是运算规则和常用值的起点不一样,数学还是一样的。
无标题无名氏No.57829192
2023-06-02(五)17:35:26
ID: rtouJd5
>>No.57828464
欧美很多国家在基础教育是不搞九九表这样的东西的,他们的基础直观是在白纸上标一个个点。他们从古希腊到康德的数学直观基础都是这套东西,甚至到恩格斯写《反杜林论》的时候从数学开始反对都是指这一套东西。用表直观更多是我们国内或者说东亚的做法。
无标题无名氏No.58499433
2023-07-08(六)01:54:26
ID: 9iJstkM
就算他们以傅立叶变换为认识基础,如果他们的世界客观上存在叠加,比如两个同相同频声源,那就会衍生出四则运算。倒是它们可能会把sint当做他们的1
无标题无名氏No.58574006
2023-07-12(三)17:08:57
ID: dFcYd8D
不知道( ゚∀。)但我认为加减法也不简单,有时候感觉四则运算就像魔法
无标题无名氏No.58522888
2023-07-09(日)16:20:40
ID: Eclhi8Y 回应
有没有数学的科普小说推荐呢?
回应有 20 篇被省略。要阅读所有回应请按下回应链接。
无标题无名氏No.58540152
2023-07-10(一)17:52:35
ID: Eclhi8Y (PO主)
>>No.58535679
谢谢!我去看看(*´∀`)
无标题无名氏No.58540214
2023-07-10(一)17:56:32
ID: 3XIUAYR
平面国算不算啊( ゚∀。)7"
无标题无名氏No.58569582
2023-07-12(三)12:05:04
ID: Eclhi8Y (PO主)
>>No.58540214
算的算的!之前就想找这本书但是忘记名字了!感谢您(=゚ω゚)=
无标题无名氏No.58570485
2023-07-12(三)13:14:35
ID: lvHX5fT
能接受漫画的话,《幻想数学大战》,我心中的数学科普神作(……)
无标题无名氏No.58571673
2023-07-12(三)14:35:24
ID: Eclhi8Y (PO主)
>>No.58570485
可以接受!感谢推荐!我去看看(*゚∇゚)
无标题无名氏No.58313124
2023-06-26(一)15:38:22
ID: 3PCHwJC 回应
正在上神经病学 联系自己看的一些知识 感觉阿尔兹海默症可能和肾精枯竭有关
脑又称髓海 由肾中先天之气营养
随着年龄增加 肾气衰竭不能支持 可能会影响脑部
个人经验 和手冲有关 肾精亏空 我时间长冲多了会有脑中发空 不能安眠( ゚∀。)
回应有 13 篇被省略。要阅读所有回应请按下回应链接。
无标题无名氏No.58478891
2023-07-06(四)22:57:27
ID: 9WBhBCx
( ゚ 3゚)倒不如说,纵欲会导致亚健康,亚健康会增加患病率
可是严格地执行健康生活,没了炸鸡和快乐水,活着还有什么意思( `д´)
无标题无名氏No.58479128
2023-07-06(四)23:09:33
ID: 0QrmmIe
>>No.58478891
健康然后变成小帅然后变成美少女和别的美少女恋爱!
无标题无名氏No.58479402
2023-07-06(四)23:24:48
ID: 9WBhBCx
>>No.58479128
( ゚ 3゚)过于健康的人生,可能会不太讨喜。我就见过一个抱怨男朋友生活太健康的。
不熬夜喝酒,基本没法以夜宵吃饭为由把他约出门。约出门了,大概率也是晨练之类的。陪他运动暴汗,妆都要花了。
无标题无名氏No.58515772
2023-07-09(日)02:01:13
ID: NwBl8vr
首先,中医所讲脏腑(在没有具体系统学过中医的人眼中)可以视为功能的集合,并不代表实质的具体器官。
其次阿尔茨海默病虽然说是脑部病变,但是主要症状出现在失忆,又表现出包括情绪、认知、行为等多方面的症状,我没有见过阿尔茨海默病的患者,不敢肯定主要原因是什么,但是从中医理论方面来讲肯定是心出问题了,同时要波及到肝脾肾。
就预防来说,手冲这件事是否推荐是要看情况的,当然古人在道德礼义这方面肯定是不推荐你手冲的,而具体到中医方面,一天一次的手冲也肯定是过度频繁了,因此不建议高强度手冲。至于手冲和阿尔茨海默病有没有强关联性……我只能说我不到啊( ゚∀。)7
无标题无名氏No.58550039
2023-07-11(二)09:14:34
ID: 3PCHwJC (PO主)
>>No.58515772
专业 可以做个课题?
无标题无名氏No.58034158
2023-06-12(一)10:44:15
ID: UkMo0dW 回应
艺术史相关,大概就是常识科普+逛各种艺术展览的心得
po只是一个学习这个专业没多长时间的学生,如果有错误的地方欢迎肥卿们指出来( ゚∀。)
回应有 24 篇被省略。要阅读所有回应请按下回应链接。
无标题无名氏No.58318326
2023-06-26(一)20:01:30
ID: UkMo0dW (PO主)
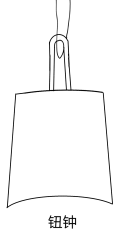
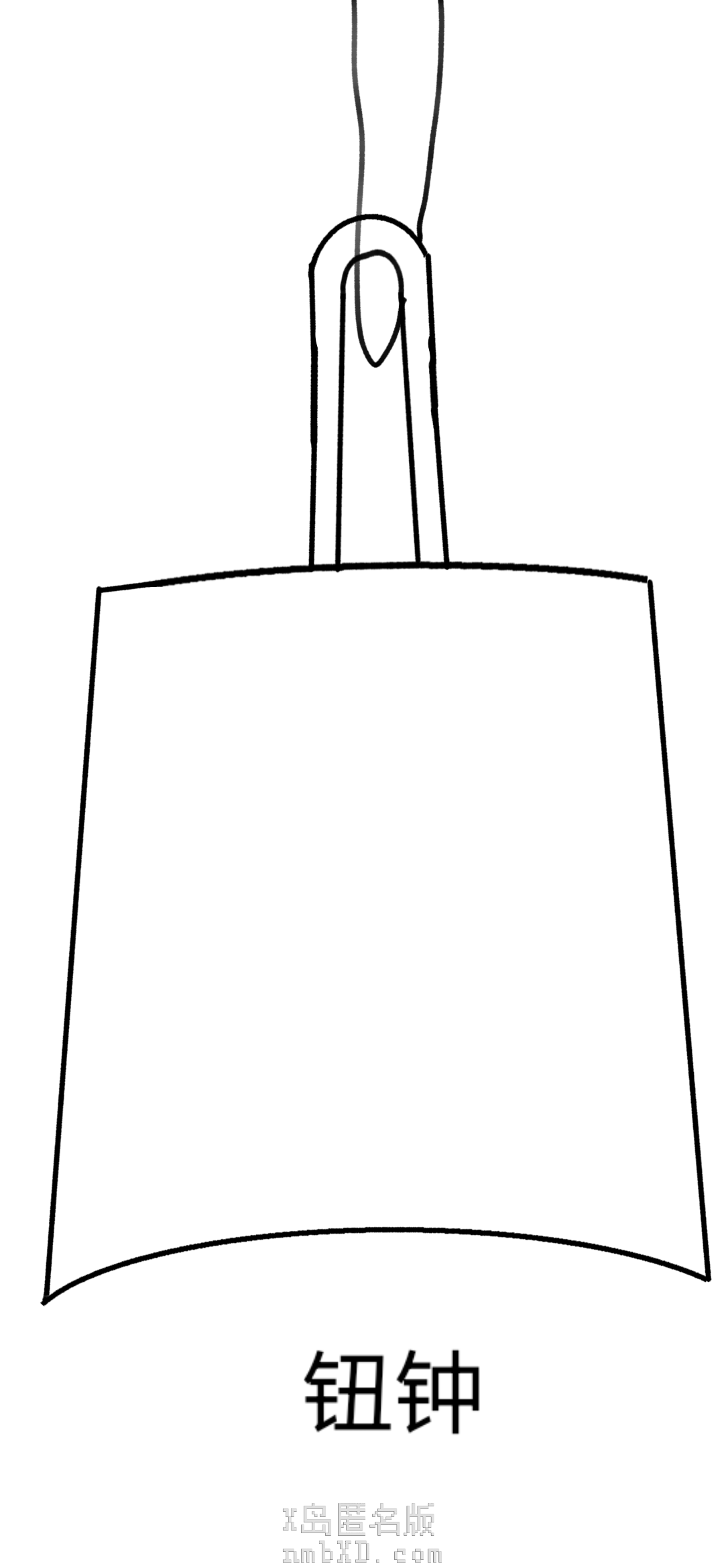
底部有弧度、直着挂的是钮钟
钮钟出现得比甬钟晚
无标题无名氏No.58318343
2023-06-26(一)20:02:21
ID: UkMo0dW (PO主)
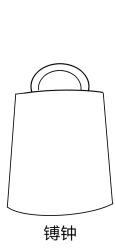
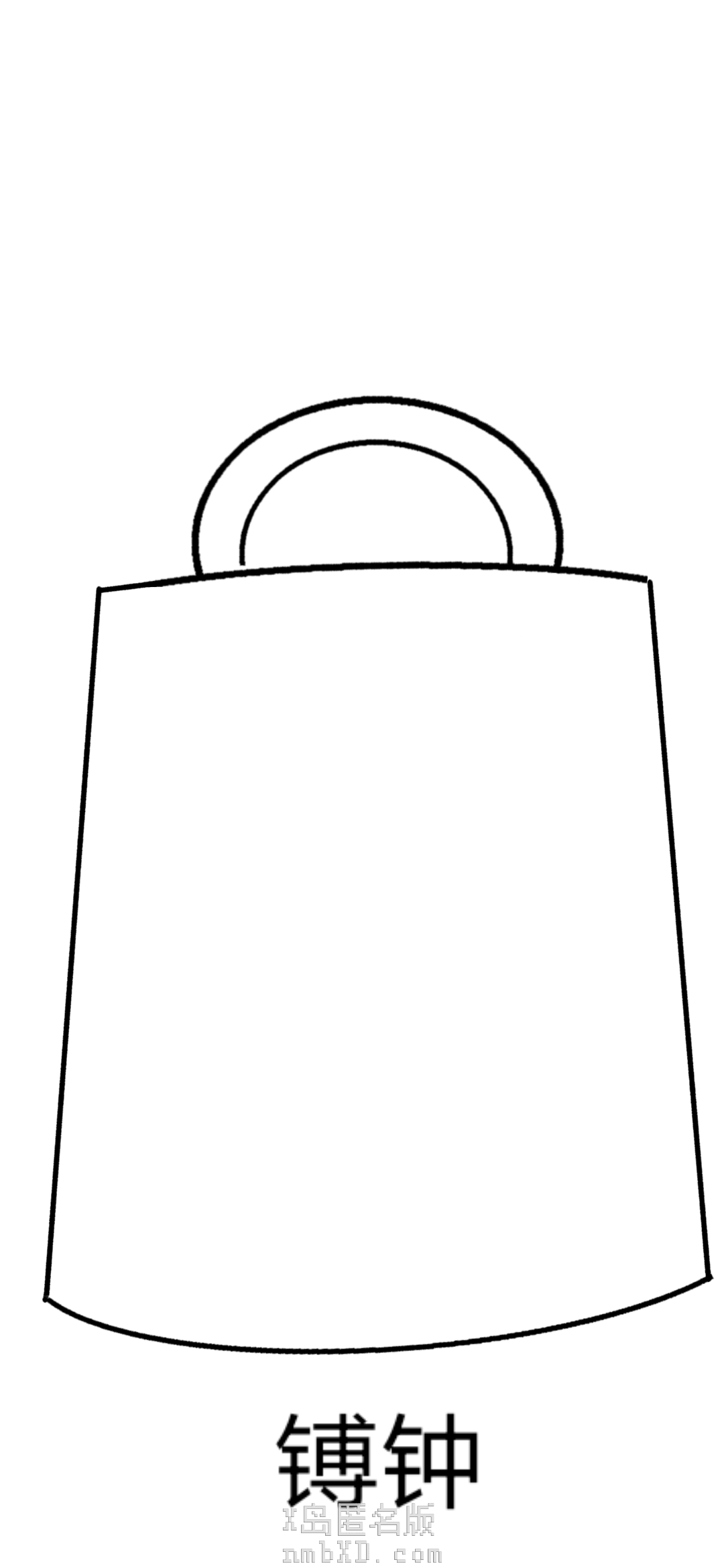
平底的是镈钟
无标题无名氏No.58362880
2023-06-29(四)21:47:59
ID: InDAgx8
椒麻鸡皮σ`∀´)
无标题无名氏No.58438053
2023-07-04(二)16:27:06
ID: BVnbh0B
jmjp
无标题无名氏No.58509591
2023-07-08(六)19:20:21
ID: Jv6FnP5
( ´∀`)摩多摩多,好有意思
无标题无名氏No.58462137
2023-07-05(三)23:16:20
ID: xpuF0Wj 回应
将逻辑和数学相关联的书有哪些?
无标题无名氏No.58462407
2023-07-05(三)23:30:39
ID: LJG0KBr
离散数学
无标题无名氏No.58462552
2023-07-05(三)23:39:20
ID: Xgbo6j9

物理逻辑+线性代数
无标题无名氏No.58463827
2023-07-06(四)01:01:27
ID: tOXRA8c
或许可以看看 3Blue1Brown 的视频
例如我刚看了他讲线代的视频,前面说的把向量视为放缩的基向量的组合,把空间视为基向量张成的空间,把线性变换看成是对基向量张成的空间的变换,这些我都知道,突然到行列式这里,跟我说行列式是单位面积缩放的比例,然后讲到特征空间啊,将坐标理解为面积(体积),来给出克拉默法则的几何解释啊,之类的,都很神奇
如果你是需要理解数学背后的直觉,就很适合
无标题无名氏No.58483899
2023-07-07(五)09:21:30
ID: OQ4nsmr
符号逻辑/数理逻辑
无标题无名氏No.58462141
2023-07-05(三)23:16:42
ID: xpuF0Wj 回应
将逻辑和数学相关联的书有哪些?
无标题无名氏No.58083177
2023-06-14(三)18:30:18
ID: xpuF0Wj 回应
为什么e会在那么多领域出现?比方说神经科学的Sigmoid函数( ゚π。)
回应有 7 篇被省略。要阅读所有回应请按下回应链接。
无标题无名氏No.58087998
2023-06-14(三)22:33:32
ID: leR52Da
谢谢,已经开始头晕了
无标题无名氏No.58096510
2023-06-15(四)12:14:52
ID: 0QrmmIe
>>No.58087880
(1+1/n)的n次方的极限嘛
无标题无名氏No.58147796
2023-06-17(六)22:55:02
ID: low6mNY
>>No.58083177
因为以e为底的指数函数有很好的性质,所以人们经常用它。
举一个其他的例子,因为多项式有很好的性质,所以人们选择有时候将函数近似为多项式来研究。
无标题无名氏No.58147902
2023-06-17(六)23:00:53
ID: hhyS5E6
其实我挺好奇欧拉公式的,可以把e和π两个无理数联系起来
无标题无名氏No.58452274
2023-07-05(三)12:42:23
ID: 6pRSsg3
>>No.58147902
欧拉公式更像是一个复数的指数形式的定义的特例,而非公式,但是在复分析里面具有广泛的应用和意义
无标题无名氏No.58414316
2023-07-03(一)07:24:09
ID: KsZQTG7 回应

来点人肉OCR和FaTP
无标题无名氏No.58414329
2023-07-03(一)07:26:34
ID: KsZQTG7 (PO主)
具体而言,关于种子植物系统演化可以在形态学、胚胎学、古植物学以及系统发育等方面找到众多相关的证据。
1.在形态特征方面,从种子蕨到现代的各类种子植物,它们的胚珠的形式有不同的来源,并发展出多种不同的类型。在种子蕨就可以找到两种不同形式的大孢子囊。一种是大孢子囊生在无叶的叉状分枝的顶端,另一种是长在孢子叶的两侧。由此演发出多种不同类型的大孢子囊:(1)银杏类的 Ginkgo、Baiera、Trichopitys 的大孢子囊(胚囊),生长在无叶的枝顶。(2)Calathospermum、Bennettites、Cordaites的多数大抱子囊直接长在简单的轴顶,并且有鳞片包着。(3) Taxales 紫杉类与 Gnetales 倪藤类的胚囊生于分枝的顶端,并托以苞叶。(4) 松柏类的胚囊生在简化的孢子叶上。(5) 苏铁与有花植物的胚囊生于大孢子叶 (心皮) 里。大孢子囊在来源和结构上的多样性这一客观事实,是和人为地把种子植物划分为裸子植物及被子植物两大类的分类法不协调的。
2.在系统发育特征方面,从种子蕨到现代高级种子植物,一直找不到统一的系统关系。它们各自出现在不同的地质年代,彼此在形态结构上又缺乏直接的联系。Cordaites科达出现在晚泥盆世,到二叠纪便趋于消亡,它和松柏类似乎不具直接的亲缘关系。Ginkgo银杏始现于晚三叠世,它和科达或种子蕨都缺乏直接的联系。它和出自早二叠世的Rhipidopsis扇叶也没有直接的联系。另一支出自晚泥盆世的 Pteridospermae种子蕨类被认为是和Bennettites本内苏铁、Cycadales苏铁类、Nilssoniales有着较直接的联系。而Pentaxylales 五柱木类、Corystospermae和 Caytoniales开通类似乎和种子蕨类不存在直接的联系。侏罗纪出现的Ephedrites麻黄及 Gnetopsis倪藤到目前为还找不到它们和哪一类有什么亲缘关系。种子植物在古生代中期开始出现,由于发生的年代久远,除了银杏和苏铁之外,大多数进入中生代中期已经消失。三叠纪是种子植物全盛时期,松柏类、银杏类、Nilssonia、苏铁类、本内苏铁类、五柱木类、盾籽类、开通类、Corystospermae等均在三叠纪获得进展,甚至有花植物也可能出现于三叠纪,可是曾几何时绝大多数种子植物到了侏罗纪便逐渐消亡,这说明大多数的种子植物在结构上多么不适应,只有少数残存了下来 (银杏、苏铁)。得到长足发展的惟有有花植物和球果类的松柏植物。由于找不到直接联系的中间类型,因此从事古生代和中生代植物区系研究的学者,多认为种子植物在起源方面是多源的。这种设想和种子植物系统发育的事实不完全一致,原始种子植物的多样性和系统上不连续,可能是它们虽有共同起源,但朝着多个不同方向发展,或者通过突变的结果,由于构造上的不完善和不适应而消亡。另一方面,那种认为种子植物之间存在着比较密切的系统关系 (Meyer,1984),而把传统的子植物类群纳入银杏、苏铁及松柏3个纲里,还把麻黄纳入银杏植物,把倪藤归入苏铁类,把种子蕨也并入银杏类的做法,都是牵强不可取的。
3.在胚胎学特征方面,种子植物从种子蕨到现代有花植物,除了上述来源不同而发展出不同的胚囊类型之外,各大类在大孢子囊(胚囊) 的结构和演化方面存在着截然不同的形式。传统的分类学把胚珠生于闭合心皮的有花植物称为被子植物,其余的种子植物归人裸子植物,这在系统发育研究工作方面曾起过积极的作用。而更多的系统学资料和研究结果显示,这种分类法存在局限性和片面性。例如银杏类的胚珠生在裸枝顶端,缺乏孕性的心皮叶。为了加强对种子的保护,银杏发展出增厚的珠被,它在受精后,珠被分化为3层的果皮,包括厚肉质的中果皮,骨质的内果皮,最外为薄层的外果皮,它比那些由心皮叶关闭起来形成子房所发育出来的真正果实,并不逊色,这是一种原始型的果实,不宜叫做种子,也不能认为银杏雌器的珠领是退化的心皮叶,不能把不同源的组织,强加给银杏。紫杉类和罗汉松类也属于这一类型。紫杉和罗汉松的珠被里有2列维管束,珠被外面有一个杯状体,一向叫它外珠被,或叫它假种皮,都是不恰当的,它和有花植物的假种皮,各有不同的起源。在杯状体内还有维管束组织(管胞)。它和银杏的珠领同一性质,它是不具心皮叶的胚珠的保护组织,以后厚肉质的珠被在受精后,继续发育也分出3层组织,加上杯状体的肉质组织构成了另一类型的果实,它不是简单的种子。这些都是种子植物在发育过程各自形成的独特的果实。在系统发育过程,这种果实是和银杏及苏铁的果实一样,在不具心皮叶这一类保护组织的条件下,发展起来的另一类型的果实。倪藤被叫做盖子植物,是因为在胚珠外侧有2层盖被,这是另一种保护种子的组织,是不具心皮叶的胚珠在形成果实过程中,特化出来的早期果实的结构。以有花植物的果实为标准,把一切早期从不同的途径发育出来的果实叫做“种子”,是不符合系统发育规律的。倪藤和百岁兰都不具颈卵器,倪藤不具管胞,在受精过程有2个精子进入胚囊,是一个雏形的双受精现象,倪藤的营养器官,包括茎和叶的各种结构都和有花植物的没有两样。
无标题无名氏No.58414330
2023-07-03(一)07:26:50
ID: KsZQTG7 (PO主)
来自具有心皮叶的胚珠,也不是全都闭合的,Caytonia开通的心皮在受精前是开放的,在受精后才闭合,这是果实的系统发育的前期产物,是未臻完善的结构,它在木兰目的Degeneria也表现了出来。苏铁类的胚珠是生在心皮叶上的,但胚珠发育过程,不依靠心皮叶关闭来保护种子,而是沿着银杏果实发育的途径,不过它比银杏进了一步,在厚肉质的珠被里,有了两列维管束,而在银杏则仅有 1列,苏铁的果实也和银杏果实的结构相同,有一层外果皮,肉质的中果皮,骨质的内果皮。
由此可见,银杏类、苏铁类、紫杉类、倪藤类乃至开通类都不宜叫裸子植物,而是具有雏形被子的种子植物。银杏由于缺乏心皮叶,由加厚的珠被、还有珠领作为保护种子的包被。苏铁类虽然有心皮叶,但在系统发育过程,只能遵循银杏的发展途径,在加厚的珠被发展出2列的维管束。紫杉类加厚的珠被也有 2 列的维管束,由于不具心皮叶,而发展为杯状体,是肉质果实的先驱。它们以这些原始的组织来对种子加强保护到了具有心皮叶的有花植物才出现了比较完善结构的子房。这种次生性结构,比起其他种子植物,保护作用要完善得多。
真正的裸子植物,可能只有具球果的松柏类植物,种子只有种衣包围着裸露在种鳞上。其他被归人裸子植物的种子植物,一类是不具孕性的心皮叶,如银杏、紫杉、倪藤等。分别以加厚的珠被、珠领、杯状体发育成果皮来保护种子;或者出现了盖被 (倪藤)作为保护组织。另一类是具有心皮叶的,随着系统发育过程也呈现出不同的变化和结构,苏铁虽有心皮叶,但胚珠的保护组织仍然沿着银杏的发育途径,以加厚的珠被形成果皮,心皮叶不具保护作用。松柏类的心皮叶沿着另一个方向发展,心皮叶托住胚珠,并未完全把它包起来,只有发展到有花植物心皮叶完全包着胚珠,起着完善的保护作用。从种子及果实的系统发育过程看,它不是沿着一条道路,而是从不同的来源各自沿着不同方向发展,从而表现了不同的进化阶段,那种把进化阶段截然分为“裸子“与“被子”的分类法,显然是人为的,为了真实反映种子与果实在系统发育的实质,建议广泛地称之为种子植物。
无标题无名氏No.58414336
2023-07-03(一)07:27:21
ID: KsZQTG7 (PO主)
有花植物的起源到目前仍不十分清楚。现存的有花植物是白垩纪或晚侏罗世的产物,原始的有花植物可以追溯到三叠纪。这些原始的有花植物在历史发展过程中不断地受到淘汰。在北美科罗拉多上三叠统的Sanmiguelia可能属于原始有花植物的代表。最近在辽西出土的,存在于1.45亿年前的辽西Archaefructus古果化石,以及最近报道在美国的大羽羊齿残骸里找到了常见于被子植物的齐墩果烷,为我们研究有花植物起源时间提供信息,但对原始有花植物的发掘与探索仍有待今后的努力。因此,原始有花植物化石的发掘,将有助于证实原始有花植物的存在,也将证实有花植物的祖先的存在和有花植物起源的年代。
发轫于三叠纪或更早的原始有花植物,延续到白垩纪获得了较完整体系的发展,原始有花植物在结构上也比现代的有花植物简单而古老,并经历了一系列的改造,才得臻于完善的结构,并在白垩纪获得迅速的发展,原始的有花植物,花的结构要比现代的真花简单得多,从小孢子叶发展为雄蕊,大孢子叶发展为雌蕊,并由不孕的孢子叶转化为花被或由苞叶转化为花被。原叶体和颈卵器的退化,出现了双受精和次生性胚乳,木质部出现导管、薄壁组织、纤维组织的转化需要有长期的改造,而这种转变和改造,不是一次完成的,只能是经过长时间的历程。因此,研究有花植物的来源及起源的时代,必须有发展的观点,不能以现代有花植物的高级结构为标准去衡量和找寻原始的有花植物。传统的研究,坚持以现代有花植物的结构为标准,否定了三叠纪以来所找到的许多有花植物化石,不承认白垩纪以前存在着有花植物,这只能把后人对有花植物起源的研究引到不可知的道路上去,因而无法解决有花植物的起源和演化的系统问题。
有花植物来自哪一类原始种子植物或蕨类植物,它在系统发育上是否直线单传的单元起源?它的起源时代如何?这些都既不清楚,也有很多争论。从有花植物的子房和胚珠的结构看来,它的祖先只能是具有异形孢子及孢子叶的原始种子植物,而不是不具孢子叶的原始种子植物。经过简单到复杂,不完善到完善的改造,才从原始种子植物发展为原始有花植物,再出现现代的有花植物。这些比较原始的有花植物在种系演替的过程中不断更新,残存的代表都是不连续的,孑遗性的。因此要寻找较原始的有花植物,不能从那些种系繁衍而连续、结构完善而复杂的代表中物色,包括木兰科在内的木兰目各个科,都不是原始有花植物的代表,这个目的各科,在地史上和化石证据都不早于白平纪。个别的可见于侏罗纪,应该在更古老、种系更残缺的代表中寻找。在华夏古陆的三叠纪、侏罗纪地层,曾发现不少有花植物的化石和花粉,甚至还有属于二叠纪的遗物。如浙江长兴二叠纪煤层里发现过双孔的有花植物花粉;四川广元的三叠纪地层发现过3沟花粉,在河南平顶山及广东的三叠纪及侏罗纪地层找到化石果,有人认为这类化石果当中,有的是木兰科的种子;在东北的燕辽平原中侏罗世地层里,找到现代的毛茛科植物的化石和大量的Sinodicotites中华缘蕨半被子植物 (?) 混生在一起,从这些事实可以相信,三叠纪已经存在着原始有花植物。现代的有花植物,都不可能是最古老的,它们是从最古老的类型通过多条途径发展出来的,属于第二阶段,或第三阶段的产物,流行的系统学把木兰目当作最原始的有花植物,并用系统树来说明全部有花植物都是从木兰目演发出来,这种单元单系的思想是和有花植物的系统发育实际不相符合的。现代生存的类型,不管是木兰目、毛目、柔荑花序类、昆栏树目、水青树目、金缕梅目、睡莲目、泽泻目等都是由不同的原始祖先演化出来的,它们之间是不连续的,而且也不是最原始的,因此彼此之间缺乏直接的亲缘关系。木兰目一向被认为是最原始的代表,但无论是木兰科、Winteraceae、Austrobaileyaceae都有某些次生特征。木兰科的种系繁衍而连续,花的结构很完善,虽然雄蕊及离生心皮是原始性状,次生木质部却具有明显的次生特征。Winteraceae具有管胞,但种系也比较发达,化石仅见于第三纪,它也和木兰科一样,花从两性分化为单性。Austrobaileyaceae具藤本习性,木质部有明显的次生
特征。
无标题无名氏No.58414340
2023-07-03(一)07:27:50
ID: KsZQTG7 (PO主)
单沟型花粉被认为是最原始的特征。但在花粉研究者当中,有人认为3沟花粉是最基本的,单沟花粉是从3沟演化来的,特别是孔型的花粉不可能是从沟型演化来的。在多心皮类,花粉都是沟型的,而在柔荑花序类,花粉基本上是孔型的。在多心皮类里,樟科及番荔枝科的花粉多数是无沟无孔的,它不可能是从单沟花粉演化出来。单子叶植物的花粉开口最富多样性,或无沟无孔,或单孔多孔,更多为单沟或多沟,还有环沟及螺旋孔等,反映出单子叶植物来源方面可能从多个方向发展出来。在浙江长兴二叠纪煤
层的双孔花粉,以及四川广元三叠纪地层的3沟花粉,说明了孔型及3沟花粉的邃古性。
关于风媒植物与虫媒植物的原始性问题,德国学派认为风媒花是原始的,多心皮学派则认为虫媒花是原始的。从种子植物系统发育过程看,无论是种子蕨、科达、银杏、开通类等都是风媒的,原始的有花植物只能从风媒的种子植物脱胎而来。毫无疑问,花粉是昆虫的重要食物来源,无论是风媒花还是虫媒花,它们的花粉都是昆虫的粮食,从昆虫出现的第一天起,或是花粉出现的第一天起,并不存在昆虫作为传粉媒介这回事。有关昆虫与植物传粉一系列所谓适应机制、适应传粉的结构,诸如分泌细胞、腺体、花形,两蕊异长异熟、昆虫口器的演化问题无疑是次生性的,认为虫媒植物先于风媒植物,风媒植物是从虫媒植物演化而来的说法,都带有明显的主观臆测性,我们认为风媒植物即使不先于虫媒植物,二者至少是齐头并进。
关于单花与花序的原始性问题,在种子植物当中,无论是开通类、科达类、银杏类、苏铁类、五柱木类,都不是单花,而且它们的花都是单性的,雄蕊与雌蕊集中在一起,组成单花的结构,是从分散到集中的过程,具有明显的次生性。因此认为单花最原始的提法,似乎缺乏足够的证据,同样地,花被的出现,有它的历史过程,为了加强保护而分化出花被,这是第一性的因素,以后才出现所谓引诱昆虫的现象,这是第二性的。
关于柔荑花序类的问题,通常认为它们是单被或无被,而且是单性的。其实在榆科、壳斗科及桦木科,甚至马尾树科都是具有2轮花被的。而榆科、荨麻科、马尾树科是具有两性花的,甚至壳斗科也有两性花的迹象。柔荑花序类基本上是具孔型的花粉,只有Quercus有沟型花粉。那种认为孔型花粉来自沟型花粉的说法是缺乏说服力的。柔荑花序类不可能从多心皮类演化而来,很有可能是和多心皮类同时由某一原始类型沿着不同的方向,分道扬镳,齐头并进。在东北燕辽地区中侏罗世地层海房沟组中发现的柔荑花序化石等支持了这一论点。诚然,柔荑花序类的木质部都不具管胞,具风媒花,花被不显眼,但它的花粉同样是昆虫(如蝇类等) 的粮食,开花期散出的臭味对昆虫具有吸引作用,人们从孔型花粉、合点受精等特征更能相信它们是从另一些非离生心皮的祖先演化出来的。
无标题无名氏No.58414342
2023-07-03(一)07:28:12
ID: KsZQTG7 (PO主)
用木兰植物一词来代表全部双子叶是不能令人满意的,至少柔荑花序类不是从木兰目演化出来。无论合瓣花类是否一个自然类群,把白花丹科和报春花科从合瓣花类移到中央子类(石竹亚纲)是不协调的。用百合植物来代表全部单子叶植物,也是牵强的,无论单子叶植物各亚纲是否具有多条发展路线和来源,百合类也不能代表泽泻目和棕榈目。
Trochodendrales昆栏树目的Trochodendraceae昆栏树科和Tetracentraceae水青树科具有3沟花粉,花被不显眼,从双被到单被,多花排成总状或头状花序,被认为是来自木兰目,并由此演化出柔荑花序。如果承认3沟花粉是有花植物的基本结构,加上木质部还保留着管胞,与现代水青树木材结构相似的化石曾发现于下侏罗统和下白垩统,它和木兰目可能不是同出自一个祖先,而且比木兰目要古老得多,虽然水青树也具有离生心皮以及染色体数目与木兰科相近 (x=19),但不能由此认为是同源的。
金缕梅科被多心皮学派认为是通向柔荑花序类的桥梁。从形态学及花粉结构看来在金缕梅科传统的6个亚科当中,Altingioideae蕈树亚科完全无花被,单性花,具柔荑花序,花粉是孔型开口,这些特征都和柔荑花序类十分一致,应该把它从金缕梅科划分出来,成立Altingiaceae蕈树科,并置于柔荑花序类里去。金缕梅科其余5个亚科,都具有花被,两性花,花粉3沟型,可能和本书所列的Trochodendridae昆栏树亚纲发生联系。
综观以上概述,本书将Spermotophyta种子植物门分为6个亚门,并拟出一个新的种子植物系统分类大纲(见第8-14页),在理论上它是一个单元多系的进化系统,在总体上可以认为种子植物是一个较完整的体系,同时,综合考虑近年来系统学研究进展合作者早些时期的观点(张宏达,1986;2000),结合本书各部分的讨论,初步设计种子植物系统分类框架见图、有花植物系统分类框架简图。